How do cells store, intraconvert, and interpretpret
information? Organisms grow and develop while their bodies are regulated
and maintained so that they can successfully reproduce in order to form
a new generation. How do all these processes function? Nucleic
acids and proteins play the roles of information molecules in cells.
Here are a few more questions worth pondering before we delve into the
information flow of cells.
What is the basic nature of information representated
by these molecules? (We will see the importance of linear sequences and
three-dimensional structures)
How is information converted from one form
to another?
How is the information selectively converted?
(In other words, how is the linear nucleotide gene sequence expressed into
animo acids?)
What kind of vocabulary can we develop to
enable us to describe and discuss information molecules?
How can we exploit experimentally what goes
on during information flow?
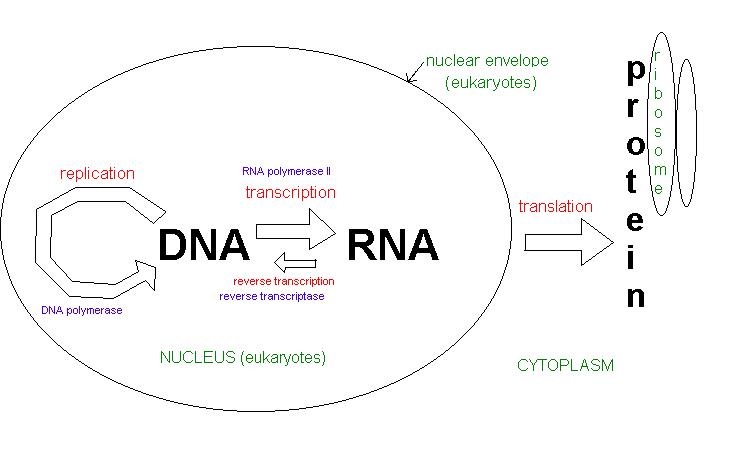
Central
Dogma
The occurrence of membrane bound organelles
for eukaryotes (more specifically, the nucleus) creates a spatial and temporal
difference. In other words, transcription occurs in the nucleus while
translation occurs in cytoplasm, and transcription/translation may proceed
at different times since they are UNCOUPLED. This uncoupling allows
for eukaryotes to increase the quality of the process by enforcing more
controls (i.e. cutting out intons) to reduce the amount of errors.
Transcription
RNA polymerase II and general transcription factors (GTFs) contribute to transcription. The primary transcript is the initial UNPROCESSED single strand molecule. Transcription begins at the 5' end where a methyl guanosine cap is created. This cap site will 1)stop degradation of the 5' RNA strand, 2)make nuclear export more efficient, and 3)make initiation of translation of mRNA more efficient as well.
Now on the 3' end, processing has 2 steps that
will "snip off" this edge of the strand at specific points by endonucleases.
1)Cleavage removal of a stretch of nucleotides at the 3' end is performed
by endonucleases and other helpers.
2)20 - 250 adenines are added by polyA polymerases
to create the "poly A tail."
The poly A tail will increase RNA stability and translatability.
Now the mRNA, consisting of only exons, must
leave the nucleus through NUCLEAR PORES.
Gene
Expression
Remember the uncoupling of transcription and translation mentioned earlier? What else can these extra steps do to increase efficiency of gene expression? Well, the many steps involved allow the cell to control various mechanisms and products at different points along the way. Proofreading, switches, delays, and other maneuvers enhance the specificity of the whole process. The numerous steps are responsible for a large amount of proteins so that MORE CAN BE DONE to improve gene expression.
Gene expression can be controlled at various stages.
1)Control at the level of transcription
There are 2 important regions of a gene, upstream promoter sequences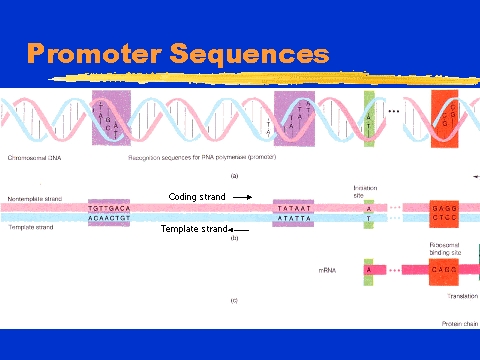 and coding sequences (where information for protein structure and function
lie). In the promotor sequences, RNA polymerase II binds to the TATA
box (initiator of transcription) by the aid of TATA-binding proteins (TBP).
DNA-bound GTFs interact with subunits bound to TBP so that DNA will loop
around. Protein-protein interactions increase transcription performance
so enhancers, which can be tens of thousands of base pairs away (and work
upstream, downstream, inverted), will promote the assembly of the basal
transcription machinery at the promoter and interact with activators and
coactivators. The CAAT and GC boxes bind transcription factors which
aid in polymerase efficiency. Transcription factors can be stimulated
(by glucocorticoids) or inhibited (by negative regulatory proteins that
bind to promoter elements) and affect the regulation of the basal level
of transcription (which is initiated by promoters). Methylation can
inactivate gene expression, and removal of acetyl groups can repress transcription.
Another way to alter proteins is to cut pieces of promoters. For
instance, blue cotton could be created.
and coding sequences (where information for protein structure and function
lie). In the promotor sequences, RNA polymerase II binds to the TATA
box (initiator of transcription) by the aid of TATA-binding proteins (TBP).
DNA-bound GTFs interact with subunits bound to TBP so that DNA will loop
around. Protein-protein interactions increase transcription performance
so enhancers, which can be tens of thousands of base pairs away (and work
upstream, downstream, inverted), will promote the assembly of the basal
transcription machinery at the promoter and interact with activators and
coactivators. The CAAT and GC boxes bind transcription factors which
aid in polymerase efficiency. Transcription factors can be stimulated
(by glucocorticoids) or inhibited (by negative regulatory proteins that
bind to promoter elements) and affect the regulation of the basal level
of transcription (which is initiated by promoters). Methylation can
inactivate gene expression, and removal of acetyl groups can repress transcription.
Another way to alter proteins is to cut pieces of promoters. For
instance, blue cotton could be created.
2)Control at the level of RNA processing
Alternative splicing allows for a gene to encode for two or more related
proteins. This is the reason parts of an intron in one cell can be
part of an exon in another cell. An example of control at the level
of RNA processing is the shifting of the poly A site to control the length
of the mRNA strand.
3)Control at the level of RNA
editing
The nucleotide base, cytosine (C), can be converted into uracil (U).
ACG codons can create start AUG codons. A strand can be shortened
when editing CAG, CAA, and CGA codons to produce stop codons.
4)Control at the level of RNA transport
If mRNA can't get out of the nucleus, it can't be translated into a protein.
As we talked about earlier, the methyl guanosine cap and poly A tail help
in RNA nuclear export and stability, respectively. Adenoviruses will
use this control to prevent host transport, but still allow for viral genes
to continue onto translation. HIV and influenza inhibit host mRNA
as well.
5)Control at the level of translation
This occurs in the cytosol. The untranslated region on the 3' side
contains information for the localization of mRNA. For example, the
bicoid gene is localized at the anterior end of the oocyte because it helps
in head and thorax development. The oskar gene is at the posterior
end for germ cell formation. Also, an unfertilized eggs will have
inhibitory proteins to mask mRNAs for developmental use. Ferritin
is a protein that grabs iron atoms. At low iron concentrations, the
repressor of ferritin's mRNA is active at the 5' end. When there
is more iron around, repressor is modified and it loses affinity for the
sequence (found in the untranslated region) for it usually binds.
The control on ferritin is lifted now so it can detoxify the cells.
6)Control of RNA stability
Half life is the time for half of the population to degrade. If proteins
live longer, there will be a larger population even though production of
each strand is accomplished at the same rate. More mRNAs will be
available to act as templates. In prokaryotes, the 5' end begins
degredation before the 3' end is even comopleted. mRNAs that are
used for short periods (i.e., cell division) will be have short lives and
those used for the production of dominant proteins (i.e., hemoglobin and
ovalbumin) live longer. Nucleotide sequences in the 3' untranslated
region have CCUCC repeats that allow for proteins to bind and stabilize
the poly A tail while poly A nucleases eat away at it. AU-rich sequences
will destabilize the tail. Note that the loss of the 3' tail will
allow for degradation to begin at the 5' end which will eventually move
to the 3' end.
Differences in gene expression can be studied by looking at the proteins created.
Proteins can act as an antigen for an antibody. The use of indicator dyes and possibly primary and secondary assays would allow for detection of specific proteins.
An easier way would be to focus on the mRNA that make the proteins because of various "tricks" that can be implemented.
Two dimensional gels can
use molecular weight and isoelectricity to identify the exact gene in question.
A metabolic label can be placed on the specimen by radioactivity.
The gene can be cut out of the gel so that one may work backwards to work
on finding the amino acid sequence.
Single stranded probes
are used to in hybridization. The known identity of the probe allows
for isolation of preferred mRNA strands.
To find a specific gene
(or all of them), a cDNA library will come in handy. Reverse transcriptase
will convert mRNA into complimentary DNA (cDNA). The double stranded
version of this can be incorporated into a cloning vected (i.e. plasmid)
and inserted into bacteria. Dilution plating will allow for each
separate colony to consist of different mRNA. This library can be
probed. The cDNA library will only contain the coding sequences and
not the upstream regulatory sequences. Another problem is that since
promoters direct the timing and extent of gene expression, their absence
allows for bias. If the cDNA in the library is not representational
of the actual amount, expression could range from "cowabunga" to just a
little bit.
To obtain the regulatory
sequences, one must start with genomic DNA. Enzyme restriction digests
will cut DNA into pieces. Single stranded versions can be inserted
into plasmids.
One genome project, total
expression monitoring, involves the representation of known nucleotide
sequences on an array created by dual hybridization of cDNA. Recently
Dr. William F. Loomis from the Center for Molecular Genetics, University
of California-San Diego, spoke at Texas Tech University . He talked
about silicon chips that contained entire cDNA libraries. Techniques
in the field of genetics and cell biology continue to facilitate research
as scientists delve into the new millennium.